
El tejido adiposo y los lípidos intracelulares son la mayor reserva energética en el cuerpo y como tal, son combustible disponible para atender las demandas energéticas necesarias en la práctica deportiva.
Esto abre una vía de gran interés para deportistas a la hora de desarrollar estrategias para maximizar la utilización de lípidos durante el ejercicio con el objetivo de mejorar el rendimiento.
Como ejemplo tenemos el consumo de dietas baja en carbohidratos (CHO) y alta en grasas, conocido como dieta cetogénica. Debido a un elevado contenido de lípidos, junto con un aporte limitado de CHO, la dieta cetogénica provoca una mayor utilización de los ácidos grasos y cetogénesis (es un proceso metabólico por el cual se producen los cuerpos cetónicos como resultado del catabolismo de los ácidos grasos)
Reprogramando el sistema metabólico
Un elemento clave que favorece el deterioro de la capacidad del rendimiento durante la práctica deportiva es la reducción de la disponibilidad de CHO asociado con la incapacidad de utilizar de forma alternativa las grasas como fuente energética. Resulta paradójico, que el declive de la función cognitiva y física están asociados a una depleción de los niveles de glucógeno y esto lo vemos en sujetos con gran cantidad de reservas energéticas en forma de grasas, pero que aparentemente no son capaces de “tirar de ellas”. Hay que tener en cuenta que la estrategia actual de poner el foco en el consumo de CHO antes y durante la práctica deportiva provoca la disminución (llegando a inhibición) de las grasas como recurso energético. Mejorar la eficiencia y acelerar la capacidad de nuestro organismo de oxidar las grasas, es posible mediante estrategias de dietas bajas en CHO que nos llevará a una situación de cetosis nutricional, acompañado del consumo alto en grasas durante un periodo de varias semanas.
En este periodo, estaremos en una situación de ceto-adaptación. Además de poder ser recurso constante de energía para el cerebro, la mayor circulación de cuerpos cetónicos, el beta hidroxibutirato, se ha demostrado que actúa como molécula de señalización capaz de alterar la expresión genética.
Hace mucho, mucho tiempo…..
Si nos vamos algo así como 2 millones de años atrás, las grasas represetaban la mayor fuente de energía para el cuerpo humano. Además de la dieta, las reservas energéticas de las que disponíamos eran fundamentalmente las grasas. Uno de los principales argumentos de los defensores de estos principios se basan precisamente en este punto, 2 millones de años de evolución deben de haber dejado mella en nuestro sistema genético.
Con la llegada de la agricultura, el ser humano adquiere la capacidad de producir enormes reservas de CHO, accediendo a un consumo constante a lo largo del año. Esto supone un incremento en la ingesta calórica, procedentes de almidones y azúcares, fundamentalmente. Se estima que las primeras sociedades agrícolas en Asia y el Creciente Fértil, comienzan hace 10.000 años (algo más tarde en sitios con peores condiciones climáticas). De esta forma, se estima que prácticamente el ser humano accede a una dieta rica en CHO en un porcentaje inferior al 1% de los últimos 2 millones de años, es decir, accedemos a CHO desde hace “un cuarto hora” (1)
En el último siglo la ciencia ha dado un salto enorme en el conocimiento del funcionamiento del metabolismo durante la inanición, reposo y ejercicio. Sobre todo, podemos ver el interés por parte de los investigadores en el apartado de dieta y actividad física, con el fin de optimizar el rendimiento deportivo. En 1939 Christensen y Hansen (2) ya se basaban en protocolos de dietas altas en CHO durante 7 días para mejorar el rendimiento físico en actividades físicas con intensidades en niveles submáximos (en lugar de dietas ricas en grasas). En 1967, Bergstrom y Hultman (3) demostraron asociación positiva entre rendimiento deportivo y concentración previa al ejercicio de glucógeno muscular. En estos años podemos encontrar varios estudios en los que la estrategia de llevar a cabo una restricción de CHO durante unos días, seguida de una carga de CHO, era interesante desde el punto de vista de maximizar las reservas de glucógeno y así mejorar la capacidad de resistencia.
Esto caló hondo y en 2002, Noakes, (y muchos otros) trabajaban bajo la idea de que sólo una dieta rica en CHO era válida con el objetivo de maximizar la capacidad del ejercicio (4).
Está aceptado a día de hoy (veamos sino los conceptos que se manejan en la Universidad) que la ingesta elevada de CHO pueden ofrecer una cierta ventaja para los deportistas, pero estos efectos no son para uniformes. Si no podemos ver la gran cantidad de deportistas que siguen un protocolo bajo en CHO y obtienen los mismos resultados (incluso en modalidades de ultra-resistencia). Esto no significa que podamos lanzarnos a recomendar abiertamente protocolos con estas distribuciones de macros (Low-CHO Diet) a todos los deportistas, pero que deben hacer reflexionar sobre que procesos metabólicos y fisiológicos están involucrados en estos casos en los que sujetos con dietas bajas en CHO llegan a obtener rendimientos igualmente válidos que deportistas que siguen un patrón establecido alto en CHO.
Ceto-adaptación
El principio de los años 80 nos dejan un buen puñado de estudios sobre la ceto-adaptación. En relación al rendimiento deportivo y la respuesta metabólica, Phinney, Bristian, Evan et al en 1983 (5) midieron la capacidad de resistencia en ciclistas antes y después de una ceto-adaptación, tras 4 semanas. La dieta era un máximo de 10 gramos al día de CHO, 15% de proteínas, resto grasas. Los valores en sangre de Beta-hidroxibutirado (BOHB) se elevaron a rangos de 1-3 mmol/L en la primera semana.
En la siguiente tabla podemos ver algunos parámetros del citado estudio,
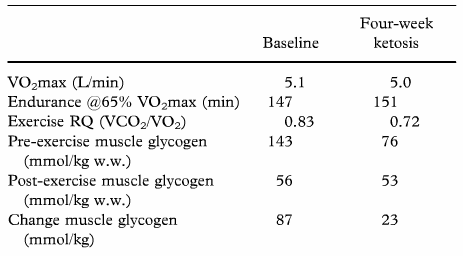
La implicación práctica del proceso de ceto-adaptación supone la capacidad por parte del organismo al uso del las grasas como principal recurso energético durante actividad en niveles submáximos, a la vez que se incrementa la capacidad de mantener las reservas de los depósitos de glucógeno en hígado y músculos en actividades de intensidad elevada. Los deportistas “ceto-adaptados” son capaces de “quemar” 90 gramos/hora procedentes de la oxidación de las grasas (810 kcal/hora) de un total de 930 kcal gastadas, en una hora de ciclismo, es decir, quedando apenas 120 kcal en una hora para “tirar” de otros sustratos como pueden ser glucosa, glicerol y aminoácidos (AA) (5).
El tiempo necesario para lograr un proceso de ceto-adaptación, podría oscilar de 3-5 días a 4-7 semanas. Es decir, al cabo de unos días de iniciar una dieta alta en grasas, y baja en CHO, la maquinaria comienza a metabolizar de forma más eficiente los lípidos como sustrato energético durante la actividad física (6)
Oxidación de las grasas durante el ejercicio físico
Intensidad
Tenemos estudios desde los años 20 de Krogh y Lindhard (7) y años 40 de Christensen y Hansen (8) en los que a través de mediciones de intercambio respiratorio, se comprobó que en situaciones de reposo y ejercicio físico con picos del 65% de Vo2max, el principal combustible energético se produce vía oxidación de las grasas.
Las fuentes energéticas procedentes de las grasas las tenemos de : i) ácidos grasos (AG) en plasma vía lipolisis en el tejido adiposo, ii) AG liberados de la hidrólisis de lipoproteínas de muy baja densidad (VLDL) y triglicéridos (TG) y AG procedentes de lipolisis de triacilglicerol localizados en gotas lipídicas en el músculo esquelético (IMTG).
Son de sobra conocidos los estudios mediante marcadores isotópicos es los que se puede comprobar lo ya comentado, el organismo tira de las grasas como combustible energético al trabajar en intensidades que van del 44% al 57% VO2max. Tenemos constancia de estos datos también al medir en pruebas de aliento basadas en sustratos marcados con carbono 13.
Estos datos no son extrapolables a cualquier tipo de activdad física. Cuando trabajamos de forma exclusiva zonas específicas, por ejemplo, en un entrenamiento de piernas, hasta intensidades del 85% se ha visto que los AG sirven como recurso energético (9). Ojo, en trabajos musculares específicos, es decir, estos datos no son extrapolables al entrenamiento por ejemplo en ciclismo. La explicación de esto podría deberse a una disminución en plasma de la concentración de AG de la hidrólisis de TG intracelulares en el tejido adiposo, lo que provocaría un descenso de la captación de AG en el músculo esquelético.
Duración
Además de la intensidad, la duración influye de forma importante en la oxidación de las grasas en la práctica de ejercicio físico. A partir de 1 hora a 1 hora y media de entrenamiento, las grasas crecen de forma exponencial en su oxidación. Por si quieres ampliar información en este sentido, tienes los estudios 10, 11 y 12.
Ayuno
Sin duda, una estrategia fisiológica para incrementar la oxidación de las grasas es el ayuno (13). Sin embargo, un ayuno prolongado acabará provocando una bajada del ratio metabólico en reposo y esto podría “contrarrestar” el efecto del ayuno respecto a las tasas de oxidación de las grasas en el entreno. De todas formas, sigo encontrando muchas sombras en este terreno y suelo ser muy prudente a la hora de recomendar protocolos basados en esta estrategia. Un ayuno prolongado provoca un incremento de los AG circulando en plasma y puede ser motivo de provocar resistencia a la insulina (14,15)
Dieta
Otra vía en la que podemos manipular los mecanismos en los que oxidamos sustratos y los usamos como fuente energética es mediante cambios en los hábitos dietéticos. Ya comenté anteriormente que en el proceso de ceto-adaptación se observa incremento en sangre de BOHB. Además, en dietas ricas en grasas durante 7 semanas, se ha visto que se ve incrementada la actividad enzimática de B hidroxi-acil CoA deshidrogenasa así como otras enzimas y proteínas involucradas en el sistema oxidativo de las grasas en el músculo esquelético.
Se calcula que los ratios de oxidación de las grasas durante el ejercicio físico van de 0.18 a 1.01 gramos por minuto (16). Estado físico de la persona, porcentaje de masa muscular, tipo de entrenamiento que se ejecuta, sexo y masa grasa, representan el 34% de la oxidación de las grasas, mientras que factores como heredabilidad y nutrición, ponderan el resto, es decir, tienen una influencia muy significativa en el proceso de oxidación de las grasas durante la práctica deportiva.
De hecho, determinados hábitos nutricionales en el pre-entreno pueden anular algunas de las adaptaciones inducidas por el entrenamiento en los genes de transcripción relacionados con el metabolismo del sustrato en cuestión. Las limitaciones más conocidas para la oxidación de las grasas serán intensidad y duración del entreno.
El consumo de carbohidratos provocará un incremento de la glucosa en sangre, y a su vez, liberación de insulina. Este paso, provocará inhibición de la lipolisis y en consecuencia, disminución de la disponibilidad de ácidos grasos no esterificados (NEFA). Esto podría llevarnos a suponer que la ingesta de alimentos con bajo índice glucémico (LGI) puede atenuar la supresión de la oxidación de las grasas, en comparación con la ingesta de alimentos con elevado índice glucémico (HGI)
En relación a lo anterior, modificando nuestro patrón dietético en pre-entreno, podremos por ejemplo, mediante la ingesta de alimentos LGI disponer de una menor cantidad de glucógeno al inicio de la actividad, y esto puede provocar el uso de una mayor cantidad de NEFA, en comparación con dietas HGI. Sería interesante en este sentido tener en cuenta el “timing” para asegurar la mayor efectividad si se pretende maximizar el uso de las grasas en el entreno.
Algunos apuntes en este sentido:
- Desayuno: podría ser un momento del día clave para consumir alimentos con LGI, al ser la primera comida tras el ayuno nocturno. Tengamos en cuenta que la exposición a NEFA durante el ayuno puede ser motivo de resistencia a la insulina.
- Alimentos ricos en fibra, presentes en muchos casos en alimentos con LGI, pueden provocar el incremento de ácidos de grasos de cadena corta (SCFA) vía fermentación en el tracto intestinal.
- El “grano entero” contiene una enorme variedad de componentes bioactivos y micronutrientes, los cuales tienen impacto e influyen en el metabolismo.
Fructosa
Siguiendo con algunos aspectos relacionados con la influencia de la dieta y el mecanismo de oxidación de las grasas en el ejercicio, tenemos el caso de la fructosa. El consumo de fructosa está asociado a : i) lipogénesis de novo, ii) acumulación de TG y iii) resistencia a la insulina. El uso de fructosa con el fin de reducir el IG de los alimentos con el fin de mejorar el uso de las grasas como recurso energético, puede no tener impacto de forma aguda, pero a largo plazo, debería tenerse en cuenta los efectos en contra.
L-carnitina
La L-carnitina juega un papel importante en el transporte de ácidos grasos de cadena larga a través de la membrana mitocondrial interna. El contenido de carnitina libre en el músculo se ve reducido en entrenamiento de alta intensidad y cuando el contenido de glucógeno es elevado. Y el contenido de carnitina libre puede limitar el ratio de oxidación de las grasas vía reducción de Carnitina palmitoil-transferasa I. Esto se ha demostrado in vitro, pero en estudios in vivo, los resultados suelen ser más conflictivos.
A la hora de recomendar la suplementación con L-carnitina con el fin de mejorar la oxidación de las grasas, los resultados nos indican que No son efectivos (17). Esto es debido a que la ingesta per se de L-carnitina es incapaz el contenido de carnitina intramuscular, incluso cuando es administrado de forma intravenosa, tampoco ser verá afectado este contenido.
Sin embargo, encontramos estudios en los que provocando una situación de hiperinsulinemia junto a hipercarnitinemia, por encima de niveles suprafisiológicos, el contenido de carnitina en músculo se vio incrementado en 14%, aspecto que a su vez, provocó un incremento del contenido de glucógeno muscular en un 30% (18). Si analizamos con detalle este estudio, comprobaremos que estas dosis fueron administradas por vía intravenosa, es decir, de difícil aplicación al deportista en general, pero abre una línea de investigación para posibles suplementos orales. He encontrado en este sentido distintos estudios con resultados no concluyentes.
Rendimiento
Debería tenerse en cuenta que el rendimiento deportivo en un proceso de iniciación a una dieta cetogénica, puede verse afectado a la baja, aún apreciándose mejoras en la oxidación del sustrato de las grasas. A partir de 4 semanas, un estrategia dietética basada en baja cantidad de CHO, mejorará no sólo el proceso de utilización de las grasas como combustible, sino que también puede presentarse mejora en la capacidad de rendimiento deportivo.
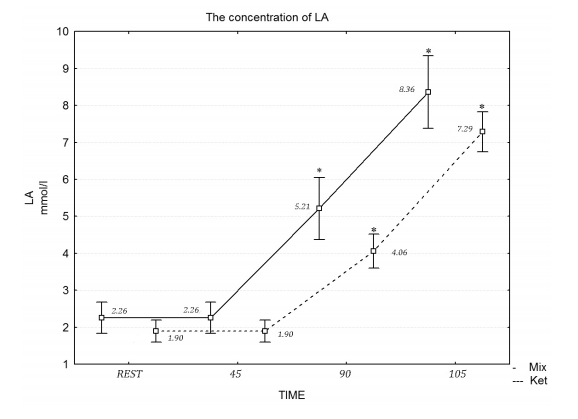
En el estudio de Zajac et al (19) compararon los efectos de una dieta cetogénica durante 4 semanas (70% grasas, 15% proteínas y 15% CHO) versus, dieta mixta (30% grasas, 20% proteínas y 50% CHO) en el rendimiento deportivo durante pruebas de ciclismo.
La dieta cetogénica aumento el gasto energético (mayor VO2) y utilización de las grasas durante la actividad moderada. Y como dato interesante, destacar que cuando la intensidad aumentó, la dieta cetogénica redujo el umbral de lactato, aspecto que supone una menor dependencia del uso de la glucosa durante el entrenamiento a intensidades elevadas.
Modificando el tipo de alimento que ingerimos en la dieta, podemos provocar o alterar el rendimiento en el entreno. El músculo esquelético ejerce una enorme influencia en el metabolismo en reposo, capacidad del entreno, regulación celular y expresión genética así como numerosas procesos relacionados con la adaptación al entreno.
Cuando nuestra dieta se basa en alta en CHO, incrementamos la cantidad de glucógeno almacenado en músculo e hígado, con la consiguiente mejora en el rendimiento deportivo, pero, de forma simultánea, incremantadmos el ratio de utilización de los CHO como combustible energético en el entreno. Esta, ha sido la base sobre las que se han basado los protocolos de nutrición deportiva durante muchos años. De igual forma, si la dieta en rica en grasas, se puede incrementar el ratio de utilización de las mismas durante el entreno.
Tenemos bastante literatura reciente en relación al impacto de dietas bajas en CHO y altas en grasas en el rendimiento ya sea en resistencia o fuerza explosiva, a la vez que tienen impacto en pérdida de peso y masa grasa. La mayoría de estudios en el caso de deportes de resistencia, indican que una dieta cetogénica mantenida durante un periodo de varias semanas, provocan que los ácidos grasos libres se convierten en la mayor fuente de combustible en el entreno y el organismo tira de los CHO de forma mucho más reducida. En el caso de estar ante un deportista que practica una modalidad de resistencia, una dieta cetogénica puede ser realmente interesante teniendo en cuenta el rendimiento (20, 21)
De forma resumida, esta estrategia dietética, nos permitiría tener una buena fuente de combustible y mantener ahorrados los depósitos de glucógeno para los momentos de mayor intensidad. En una prueba de una duración de entre 1 hora a 4 horas (resistencia) sería a tener en cuenta.
Tenemos que resaltar que todo lo mencionado, casi como siempre, resultatá efectivo en personas entrenadas. La capacidad de oxidación de las grasas es mayor en sujetos con buena forma física (20). En deportes con una duración mayor a 1 hora, la intensidad varía de baja a máxima. El papel de las grasas en el metabolismo energético durante este tipo de deportes irán de la mano de la intensidad a la que desarrollamos el ejercicio.
La persona con un cierto desarrollo muscular presenta una mayor densidad mitocondrial y capilar, aspecto que le permite oxidar grasas en mayor proporción que sujetos sedentarios no entrenados. Este ahorro de glucógeno, le permite a estas personas prolongar la sesión y evitar una depleción del glucógeno, y por tanto, retrasar la aparición de fatiga.
Otro aspecto interesante en relación a los individuos entrenados es que ven incrementada la actividad de la hormona sensitiva a la lipasa y un descenso en la secreción de insulina, tanto en reposo, como haciendo deporte.
Las personas entrenadas son capaces de “entregar” una mayor cantidad de oxígeno y sangre al músculo. Además, producen menos lactato con la misma carga debibo a un umbral más elavado de lactato. Todos estos cambios adaptativos, permiten oxidar grasas. Por todo lo mencionado, en deportes de resistencia, una estrategia basada en un protocolo alto en grasas y bajo en CHO debería al menos considerarse como opción muy interesante.
Para finalizar, las sombras de este protocolo
Hasta ahora, hemos informado de la cara A del protocolo, pero como en todo, hay que poner en la balance posibles inconvenientes.
NO para deportes de alta intensidad
El principal inconveniente es que por unidad de tiempo, una mayor cantidad de ATP puede ser generado a partir de CHO, en comparación con las grasas. El máximo ratio de resíntesis de ATP es apromixadamente 0.40 moL/min, mientras que en el caso de la degradación del glucógeno, de 1.0 a 2.0 mol de ATP por minuto.
En el caso de entrenamiento de alta intensidad, pensemos que será de elección para nuestro organismo…. Esto limitará el uso de las grasas en disciplinas que requieran trabajos a intensidades elevadas. Además, el entreno de alta intensidad suprime la lipolisis, y en consecuencia, reducción de la disponibilidad de ácidos grasos en el músculo.
Un incremento del ratio de glucólisis y producción de lactato durante el entreno de alta intensidad impedirá la oxidación de las grasas debido a una reducción de ácidos grasas de cadena larga en la mitocondria.
Cetosis
A pesar de todos los beneficios mencionados, una de las posibles consecuencias que deben tenerse en cuenta, es la cetosis. Además, relacionado con el deporte, podemos encontrarnos con deshidratación, hipoglucemia e incremento de riesgo problema renal.
Acidosis, pérdida de peso, crecimiento inadecuado, hiperlipidemia, deficiencia en algunas vitaminas y elementos traza como zinc y selenio, hiperuricemia y anemia, pueden también aparecer.
Por todo lo mencionado, aconsejo controlar este tipo de protocolos con profesionales y medios adecuados que permitan monitorizar la evolución y conocer el estado de parámetros clave.
Abreviaturas
CHO = carbohidratos
BOHB = betahidroxibutirado
AA = aminoácidos
AG = ácidos grasos
VLDL = lipoproteínas de muy baja densidad
TG = triglicéridos
IMTG = almacenamiento lipídico en músculo esquelético
NEFA = ácidos grasos no esterificados
LGI = bajo índice glucémico
HGI = elevado índice glucémico
SCFA = ácidos grasos de cadena corta
Fuentes,
(1) Orr, J. B., & Gilks, J. L. (1931). The physique and health of two African tribes. London: His Majesty’s Stationary office
(2) Christensen, E. H., & Hansen, O. (1939). Arbeitsfähigkeit und Ernährung. Skandinavisches Archiv Für Physiologie, 81, 160–171. doi:10.1111/j.1748-1716.1939.tb01320.x
(3) J Bergström; E Hultman. Muscle glycogen synthesis after exercise: an enhancing factor localized to the muscle cells in man. Nature. 1966, PMID: 5954569
(4) Noakes, T. D. (2002). Lore of running (4th ed.). Champaign, IL: Human kinetics.
(5) Phinney, S. D., Bistrian, B. R., Evans, W. J., Gervino, E., & Blackburn, G. L. (1983). The human metabolic response to chronic ketosis without caloric restriction: Preservation of submaximal exercise capability with reduced carbohydrate oxidation. Metabolism, 32, 769–776. doi:10.1016/0026-0495(83)90106-3
(6) B. Kiens, T. J. Alsted and J. Jeppesen. Factors regulating fat oxidation in human skeletal muscle. Obesity reviews 2011 International Association for the Study of Obesity 12, 852–858
(7) Krogh A, Lindhard L. The relative volume of fat and carbohydrate as sources of muscle energy. With appendics on the correlation between standard metabolism and the respiratory quotient during rest and work. Biochem J 1920; 14: 290–363
(8) Christensen EH, Hansen O. Respiratorisher Quotient und O2 Aufname. Skandinavisches Archiv fur Physiologie 1939; 81: 180–189.
(9) Helge JW, Stallknecht B, Richter EA, Galbo H, Kiens B. Muscle metabolism during graded quadriceps exercise in man. J Physiol 2007; 581(Pt 3): 1247–1258
(10) Roepstorff C, Steffensen CH, Madsen M, Stallknecht B, Kanstrup I-L, Richter EA, Kiens B. Gender differences in substrate utilization during submaximal exercise in endurance-trained subjects. Am J Physiol Endocrinol Metab 2002; 282: E435– E447.
(11) Coyle EF, Hagberg JM, Hurley BF, Martin WH, Ehsani AA, Holloszy JO. Carbohydrate feeding during prolonged strenuous exercise can delay fatigue. J Appl Physiol 1983; 55: 230–235.
(12) Kirwan JP, Cyr-Campbell D, Campbell WW, Scheiber J, Evans WJ. Effects of moderate and high glycemic index meals on metabolism and exercise performance. Metabolism 2001; 50: 849–855
(13) Cahill GF, Jr. Fuel metabolism in starvation. Annu Rev Nutr 2006; 26: 1–22
(14) Hoeks J, van Herpen NA, Mensink M, Moonen-Kornips E, van Beurden D, Hesselink MK, Schrauwen P. Prolonged fasting identifies skeletal muscle mitochondrial dysfunction as consequence
rather than cause of human insulin resistance. Diabetes 2010; 59: 2117–2125
(15) Hoeg LD, Sjoberg KA, Jeppesen J, Jensen TE, Frosig C, Birk JB, Bisiani B, Hiscock N, Pilegaard H, Wojtaszewski JF, Richter EA, Kiens B. Lipid-induced insulin resistance affects women less
than men and is not accompanied by inflammation or impaired proximal insulin signaling. Diabetes 2011; 60: 64–73
(16) Venables MC, Achten J & Jeukendrup AE (2005) Determinants of fat oxidation during exercise in healthy men and women: a cross-sectional study. J Appl Physiol 98, 160–167
(17) Hawley JA, Brouns F & Jeukendrup A (1998) Strategies to enhance fat utilisation during exercise. Sports Med 25, 241–257
(18) Stephens FB, Constantin-Teodosiu D, Laithwaite D, et al. (2006) An acute increase in skeletal muscle carnitine content alters fuel metabolism in resting human skeletal muscle. J Clin Endocrinol Metab 91, 5013–5018
(19) Adam Zajac , Stanisław Poprzecki , Adam Maszczyk , Miłosz Czuba , Małgorzata Michalczyk and Grzegorz Zydek; ; The Effects of a Ketogenic Diet on Exercise Metabolism and Physical Performance in Off-Road Cyclists; Nutrients 2014, 6(7), 2493-2508
(20) Pitsiladis, Y.; Smith, I.; Maughan, R. Increased fat availability enhances the capacity of trained individuals to perform prolonged exercise. Med. Sci. Sports Exerc. 1999, 31, 1570–1579.
(21) Yeo, W.K.; Carey, B.A.; Spriet, L.; Haeley, L.; John, A. Fat adaptation in well-trained athletes: Effects on cell metabolism. Appl. Physiol. Nutr. Metab. 2011, 36, 12